Okay, here come the doodles based on Skulachev's paper
Membrane-linked energy buffering as the biological function of Na+/K+ gradient
This is the scenario in ultra modern bacteria, the pinnacle of about 4 billion years of evolution. The membrane is tight to all significant ions at reasonable temperatures and concentration gradients. In this set of pictures the proton population represented within the red circle is holding a membrane voltage of 180mV, as per usual:

The trans-membrane potential from the pumped protons is stable while ever the pumping and the consumption of protons is balanced. The problem is that it doesn't need many protons to generate that 180mV. Pumping any more than basic needs generates too great a membrane voltage. The converse is that it doesn't take much excess proton consumption to collapse the potential. So you need a buffer which does not waste the energy used to pump.
If a bacterium suddenly increases proton pumping by eating some glucose we have this problem of a spike in membrane voltage:

We can get around this by allowing a positive ion to travel in the opposite direction. This will stop the rising membrane potential as the ion uses the membrane potential to enter the cell against a concentration gradient. It uses an ion-specific channel, in this case for potassium. This process is electrophoresis down the electrical gradient, against a concentration gradient, powered by the electrical component rather than the pH component of the rising proton gradient:

The number of K+ ions matches the excess protons pumped. The electrical potential is thus maintained at 180mV at the "cost" or "benefit" (semantics here!) of K+ entering the cell. But there is a problem in that the more protons pumped and the more K+ entering the cell, the higher the pH of the intracellular medium becomes. That K+ pool is actually tied to the OH- left behind by pumping out H+. Caustic potash...

This is not good for metabolic processes. But it is easily surmounted using a 1:1 ratio Na+/H+ (electro-neutral) antiporter to get some protons back in to the cell to offset the excess OH-
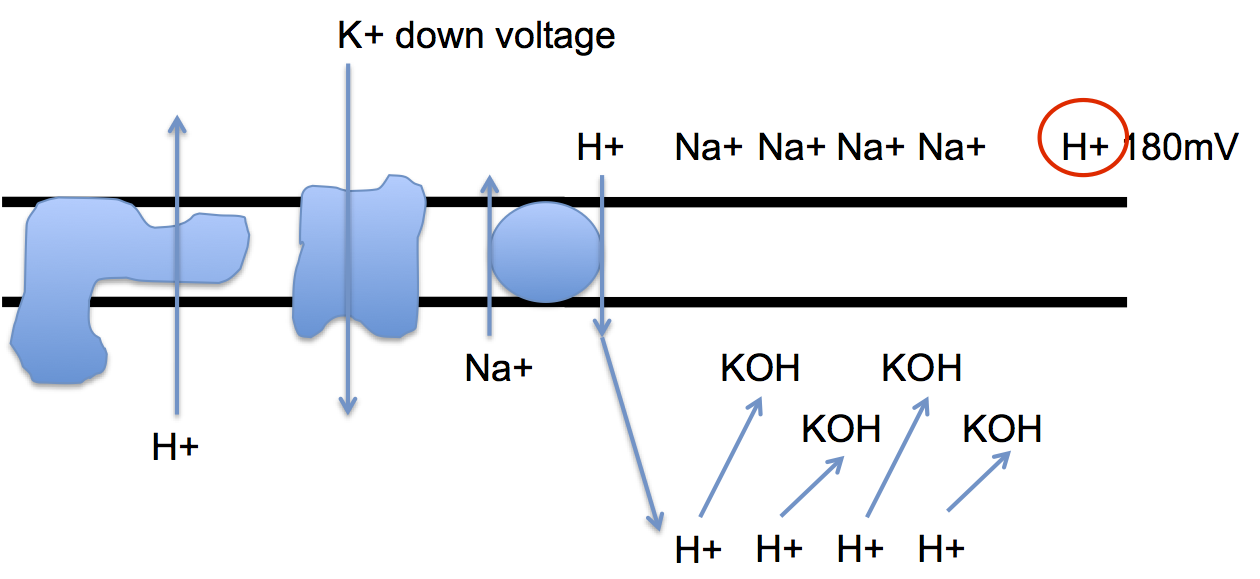
while still maintaining an electrical gradient of 180mV using H+, keeping an electro-neutral Na+/K+ gradient as an energy store:

Obviously the Na+/H+ antiporter is being driven by the pH component of the proton gradient. It's neat how evolution has separated out the pH and electrical components of a proton gradient!
The whole system is fully reversible so if there is a sudden drop in proton pumping the transmembrane Na+/K+ gradient can be reconverted to a proton gradient to "buffer" changes in proton translocation. This seems to be how modern, proton pumping bacteria with superbly proton tight membranes work. In E coli the ion channel and antiporter are ATP gated.
That's how Skulachev looked at modern bacteria in 1978.
I'm now going to wander off on my own and speculate about LUCA with a proton leaky but Na+/K+ tight membrane. This is just me from here onwards:
Let's have a think about LUCA, with a cell membrane which is tight to Na+, and probably K+ too, but highly leaky to both protons and hydroxyl ions. Metabolism is based on Na+ pumping and a Na+ specific ATP synthase. The initial Na+/H+ antiporter (from the Life series) is gone as a source of Na+ gradient as soon as LUCA leaves the alkaline hydrothermal vents.
I like the idea that LUCA used a pyrophosphatase to pump Na+ but with any Na+ pump we have the same problem as in modern bacteria: You can only store a small amount of energy as a 180mV Na+ gradient, as per H+ above:

But excess Na+ pumping can be easily be accommodated by K+ electrophoresis:

There is no need for the Na+/H+ antiporter in this scenario because there is no pH change associated with pumping Na+ ions, so all we need is the ion specific channel for K+.
This sets up a non-electrical energy store which is "accessible" to form an electrical gradient when primary Na+ pumping is low.
The buffer automatically implies the generation of a raised intracellular K+. We have here, based on a tiny step beyond Skulachev's ideas, a place within LUCA which is potassium rich. It's simply produced to buffer changes in ion pumping by the primary Na+ pump (or usage by ATP synthase) across relatively primitive membranes. And driving intracellular K+ higher is an indicator to the cell that there is excess of energy available, which should select for increased enzyme activity based on rising intracellular K+ concentration. Many of the "core" LUCA enzymes do indeed use K+ as a cofactor to function optimally.
Summary: Cells become islands of raised K+ ion concentration when more than basal a level of energy is supplied. Remember that for our later discussion about Mulkidjanian's ideas on the origin of life on Earth.
Peter

4 comments:
Thank you for this very clear exposé.
As someone who has a serious imbalance of Na+ and K+ (late-onset pseudohypoaldosteronism type 1) my body keeps me well aware of the carb intake and K+ levels dynamics... actuely so.
Did you know resting too long at night after having had carbs at dinner can cause hyperkalemic paralysis the next morning ? It's a lot less fun than it sounds. Maybe having too much energy lying around and not any mean to do something useful with it is not a good thing, metabolically.
In situations where the organism is depleted of Na+ and has an excess of K+, could the mechanism you describe for compensating the alkalization of the intracellular medium fail ? I know I always get metabolic acidosis after prolonged efforts, it could be developping in the intercellular level. Going alkaline inside, and acid outside, hmm...
In my experience, it also causes the worst kind of headache one could ever have: astrocytic swelling. Do you think there is any chance this inner/outer pH imbalance could be made worse in glucose-dependant cells, specifically ? What could be the osmotic consequences ?
Hi,
Would these proton pumping mechanisms have anything to say about the weird 'alkaline diet'?, or possibly eating foods that are very high in potassium like fruits. Avocado of course for keto :)
Thank you, Sam
So our cells use sort of a potassium battery .
https://en.wikipedia.org/wiki/Potassium-ion_battery
What happens after you run out of potassium?
This appears to stabilize things when food appears and disappears - covers the feedback zero-crossing glitch. Without it - I think there would be ROS signaling noise.
,.,.
I'm also remembering the some connection of potassium with sore muscles...
Hi Jesrad,
Ouch. Skulachev does go on to talk about mammalian cells in the later section of the paper but I’ve not thought it through to the level of looking at aldosterone dyscrasias. I have to agree the recreational potential seems limited!!!!
Samuel and karl, the body controls pH very, very tightly so I’m not one for alkaline diet ideas. Equally, unless you are on an ACE inhibitor (or have Addisons or jesrad’s problems!) you are going to off load excess K+ down the loo. Now, dealing with a deficiency of K+ is a much harder problem to sort out. But, on the whole organism basis, fat doesn’t use the insulin/K+ system in the same way as glucose…
Peter
Post a Comment